



Содержание
Перейти к:
 А. А. Венедиктов,
К. С. Покидова,
Д. М. Оганесян,
В. В. Голоборщева,
Г. А. Пьявченко,
С. Л. Кузнецов
А. А. Венедиктов,
К. С. Покидова,
Д. М. Оганесян,
В. В. Голоборщева,
Г. А. Пьявченко,
С. Л. Кузнецов https://doi.org/10.47093/2218-7332.2024.15.3.48-57
Перейти к:
Цель. Изучить экспрессию глиального фибриллярного кислого белка (glial fibrillary acidic protein, GFAP) и ионизированной кальций-связывающей адаптерной молекулы 1 (ionized calcium-binding adapter molecule 1, Iba1) в нейроглии цингулярной и ретросплениальной коры больших полушарий головного мозга у мышей на 5-е сутки после внутрибрюшинного (в/б) введения бактериального липополисахарида (ЛПС) в дозе, не вызывающей воспаление нервной ткани.
Материалы и методы. Работа выполнена на 10 самках мышей линии C57BL6 возрастом 90 ± 3 суток, массой 28,0 ± 3,2 г. На протяжении 4 дней в одно и то же время животным группы 1 вводили в/б физиологический раствор (NaCl 0,9%), группы 2 – эндотоксин ЛПС E. coli в дозе 1 мг/кг/сутки. На пятые сутки мышей выводили из эксперимента путем декапитации после введения ксилазина и тилетамина/золазепама, после чего изготавливали гистологические препараты цингулярной и ретросплениальной коры, окрашивали антителами к GFAP и Iba1. При помощи программного обеспечения QuPath подсчитывали количество: (1) GFAP-положительных клеток, (2) участки Iba1-положительной цитоплазмы. Для сравнения групп использовали U-критерий Манна – Уитни.
Результаты. Количество GFAP-положительных клеток после введения ЛПС в группе 2 было статистически значимо больше, чем в группе 1, и составило 22,5 (8,0; 32,0) vs. 9,0 (4,3; 17,0), p = 0,0038. Количество Iba1-положительных участков цитоплазмы также было статистически значимо больше в группе 2: 207,5 (154,8; 295,8) vs. 128 (89,3; 165,5), p = 0,014. В обеих группах не выявлено признаков воспаления: повышенного кровенаполнения сосудов и кровоизлияний, периваскулярного отека или миграции лейкоцитов.
Заключение. Доза ЛПС 1 мг/кг/сутки, вводимая в/б мышам в течение 4 дней, позволяет оценить изменения в центральной нервной системе со стороны глии при повреждении, не сопровождающемся развитием воспалительной реакции: в цингулярной и ретросплениальной коре увеличивается количество астроцитов с положительной реакцией антител к GFAP, а также макрофагов с экспрессией белка Iba1.
Астроциты и клетки микроглии центральной нервной системы (ЦНС) в первую очередь подвергаются изменениям при воздействии проникающих через гематоэнцефалический барьер (ГЭБ) и повреждающих структуру нервной ткани химических соединений [1][2]. В то же время изменения в астроцитах и клетках микроглии характерны для нейродегенеративных расстройств [1], нарушений памяти и обучения [3], старения [4].
Воздействия различных инфекционных и неинфекционных агентов на нервную ткань, приводящие к изменениям в популяциях макро- и микроглии, позволяют моделировать повреждение ЦНС и изучать его особенности. Исследования в этой области актуальны в связи с ростом нейродегенеративных заболеваний ЦНС и их влиянием на эпидемиологические и социально-экономические показатели благополучия населения, что побуждает искать терапевтические подходы и модели их апробации [5][6].
Отмечено, что внутрибрюшинное (в/б) введение грызунам несептических доз бактериального липополисахарида (ЛПС) может быть использовано для оценки повреждения нейроглии [7]. После первого введения ЛПС астроциты и клетки микроглии реагируют выделением различных цитокинов, а спустя несколько суток в них становятся заметны структурные изменения. Это продемонстрировано при исследовании морфологии астроцитов и клеток микроглии в префронтальной коре больших полушарий головного мозга и гиппокампе после введения мышам высоких доз ЛПС однократно или невысоких доз ЛПС на протяжении нескольких дней [2].
В качестве направлений дальнейших исследований в ряде обзоров было предложено изучить изменения в астроцитах и микроглии при введении мышам несептических доз ЛПС в течение нескольких дней. Эти клетки цингулярной и ретросплениальной коры больших полушарий первыми затрагиваются при возрастных заболеваниях ЦНС, нарушениях памяти и аффекта [8][9].
Одним из ключевых маркеров для оценки состояния астроцитов является глиальный фибриллярный кислый белок (glial fibrillary acidic protein, GFAP). Он входит в состав промежуточных филаментов в астроцитах и обладает высокой специфичностью к ним. Количество GFAP в цитоплазме клеток глии увеличивается при повреждении нервной ткани и при старении [10]. Применение другого маркера, ионизированной кальций-связывающей адаптерной молекулы 1 (ionized calcium-binding adapter molecule 1, Iba1), позволяет выявить клетки микроглии в ЦНС [11]. Основная роль этого белка состоит в стабильном связывании нитей актина в пучки, поэтому его экспрессия в клетке нарастает при переходе микроглии из покоящегося состояния в активированное [12]. Следовательно, изменения экспрессии GFAP и Iba1 служат наиболее приемлемым показателем воздействия ЛПС на клетки астро- и микроглии [13].
Цель исследования: изучить экспрессию GFAP и Iba1 в нейроглии цингулярной и ретросплениальной коры больших полушарий головного мозга у мышей на 5-е сутки после в/б введения бактериального ЛПС в дозе, не вызывающей воспаление нервной ткани.
Исследование выполнено на самках мышей линии С57BL/6 (n = 10), полученных из филиала «Столбовая» ФГБУН «Научный центр биомедицинских технологий» ФМБА России. Возраст животных на начало эксперимента составлял 90 ± 3 суток, средняя масса 28,0 ± 3,2 г. Выбор пола грызунов обусловлен данными литературы о более стойкой экспрессии толл-подобного рецептора 4 в нейроглии у самок, тогда как именно он опосредует ЛПС-сигнализацию [14]. Возраст мышей удовлетворяет двум принципам: избежать связанной со старением перестройки нервной ткани и не допустить искажения, которое у более молодых особей возможно из-за незавершенного прунинга и образования синапсов. Минимальное количество животных для разработки модели отобрано согласно концепции 3R и стандартным правилам для дизайна экспериментов на грызунах [15].
Мышей содержали в лабораторном виварии в условиях постоянной комнатной температуры 20–22 °C и относительной влажности 50–60% с доступом к чистой воде и гранулированному корму ad libitum. Перед экспериментом животные прошли двухнедельный карантин и осмотр ветеринарным врачом. После этого грызунов рандомизировали методом случайных чисел на две равные группы и с этого момента содержали мышей каждой группы в отдельной клетке.
Животным группы 1 (n = 5) вводили 0,1 мл физиологического раствора (NaCl 0,9%) в/б, а мышам группы 2 (n = 5) ЛПС 1 мг/кг E. coli O111:B4 (партия 091M4031V, Sigma-Aldrich, США), разведенного физиологическим раствором до 0,1 мл. По данным литературы, экспрессия GFAP и Iba1 нарастает после введения мышам ЛПС 0,5–2 мг/кг в/б [2][7]. На основе анализа этих методик для эксперимента выбрана усредненная доза ЛПС в 1 мг/кг. Инъекции выполняли в одно и то же время суток в течение последовательных 4 дней согласно общепринятой методике [16]. Выведение животных из эксперимента осуществляли путем декапитации с предварительным введением 5 мг/кг ксилазина гидрохлорида (Interchemie, Нидерланды) и 40 мг/кг тилетамина/золазепама (Vibrac, Франция) (рис. 1).
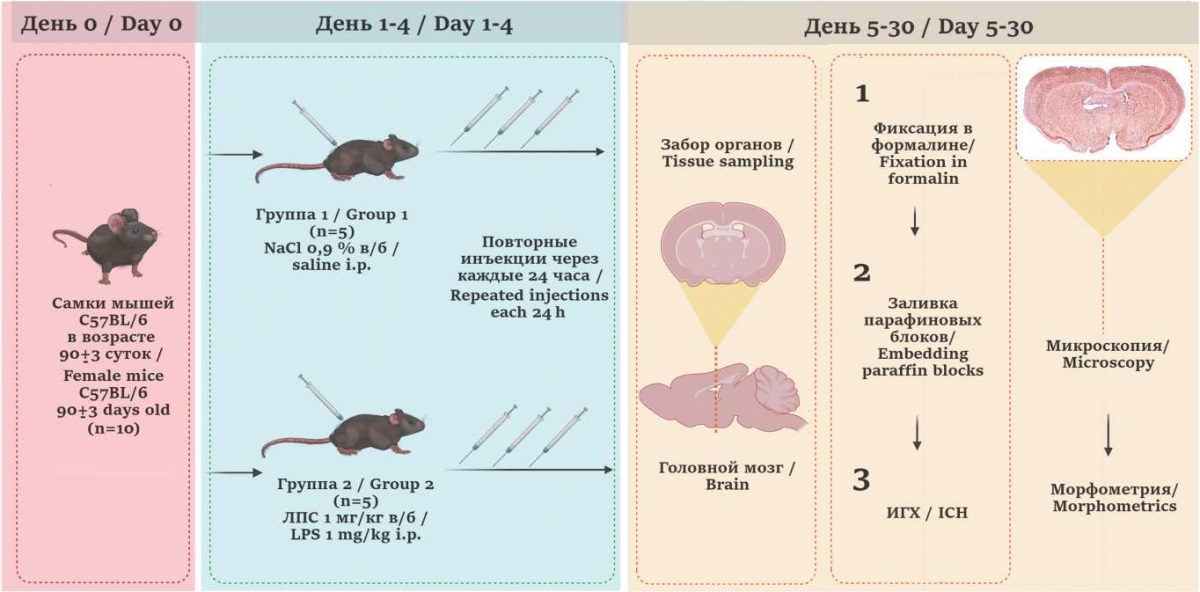
РИС. 1. Схема эксперимента.
Примечание: в/б – внутрибрюшинно; ЛПС – бактериальный липополисахарид; GFAP – glial fibrillary acidic protein, глиальный фибриллярный кислый белок; ИГХ – иммуногистохимическое исследование; Iba1 – ionized calcium-binding adapter molecule 1, ионизированная кальций-связывающая адаптерная молекула 1.
FIG. 1. Experimental design.
Note: i.p. – intraperitoneally; LPS – bacterial lipopolysaccharide; IHC – Immunohistochemistry; GFAP – glial fibrillary acidic protein; Iba1 – ionized calcium-binding adapter molecule 1.
Забор органов и тканей осуществляли по следующей методике. Ножницами разрезали кожу на уровне затылка и делали продольный разрез по направлению к носу. После отделения кожи рассекали череп поперечно на уровне носовой перегородки и далее продолжали разрезать затылочную и теменную кости так, чтобы ножницы не слишком глубоко проникали в носовую полость. Удалив череп, извлекали головной мозг. Фиксацию проводили в 10% нейтральном забуференном формалине (ООО «ЭргоПродакшн», Россия). После этого выполняли стандартную проводку через изопропиловый спирт возрастающей концентрации и заливку в парафиновые блоки. На ручном ротационном микротоме HM 325 (Thermo Fisher Scientific, США) изготавливали фронтальные срезы головного мозга толщиной 5 мкм, переносили на предметные стекла с адгезивным покрытием силан (ООО «Минимед», Россия) и подсушивали в термостате при температуре 37 °C в течение 25 суток.
Перед окрашиванием срезы головного мозга выдерживали в депарафинизирующем и демаскировочном фосфатном растворе (партия XF05RT4N9592) (Elabscience, США), разведенном дистиллированной водой 1:20, с нагревом в течение 20 мин. После этого промывали каждый срез фосфатно-солевым буферным раствором (phosphate-buffered saline, PBS) (ООО «ЭКО-СЕРВИС», Россия) трижды по 2 мин и во избежание собственной пероксидазной активности тканей инкубировали с 3% перекисью водорода в течение 20 мин. Далее, после вторичной промывки PBS трижды по 2 мин, наносили на срезы бычий сывороточный альбумин на 20 мин. После удаления альбумина препараты инкубировали с первичными моноклональными антителами к GFAP1 в разведении 1:500 и поликлональными антителами к Iba12 в разведении 1:200. Сразу же после этого каждый срез промывали PBS трижды по 2 мин и наносили вторичные поликлональные антитела кролика с пероксидазой хрена3, за чем следовала получасовая инкубация и промывка PBS раствором. В качестве хромогена использовали 3,3’-диаминобензидин (CDH, Индия), излишки которого смывали дистиллированной водой и докрашивали препараты гематоксилином Карацци (ГК «Биовитрум», Россия) с заключением в монтирующую среду.
Морфологические изменения изучали при помощи микроскопа Axio Imager.A1 с камерой Axiocam 305 color и программного обеспечения Zen 3.3 (все – производство Zeiss, Германия). Гистологическую оценку нормальной цито- и миелоархитектоники проводили на серийных срезах изокортекса. На микрофотографиях больших полушарий определяли зоны цингулярной и ретросплениальной коры при помощи стереотаксического атласа G. Paxinos и K.B.J. Franklin [17]: для подсчета были отобраны 12 корональных срезов между уровнями брегмы -0,2 и -0,3 на каждую группу. Для подсчета макрофагов снимки, полученные при увеличении объектива ×40 и фотографировании камерой Axiocam 305 color, обрабатывали с помощью программного обеспечения с открытым кодом QuPath 0.5.0 (Queen’s University Belfast, Великобритания)4 [18] в режиме обнаружения 3,3’-диаминобензидин положительных структур в одном поле зрения.
Проведенное иммуногистохимическое (ИГХ) исследование показало неравномерное распределение белка Iba1 по цитоплазме клеток – обнаружены клетки как с множеством зон с Iba1-положительным окрашиванием, так и с единичными структурами. Поэтому в качестве показателя активации клеток микроглии интерпретировали участки Iba1-положительной цитоплазмы, а не сами клетки.
Маркер GFAP, напротив, распределялся в цитоплазме клеток равномерно, поэтому для количественной оценки его экспрессии учитывали число окрашенных клеток. Подсчет количества GFAP-положительных клеток производили в программном обеспечении Zen 3.3 по снимкам, сделанным при увеличении объектива ×10 и фотографировании камерой Axiocam 305 color, в одном поле зрения.
Несмотря на то что олигодендроциты также относятся к нейроглии, их оценка в рамках настоящей работы представляется нецелесообразной. Поскольку олигодендроциты не входят в состав ГЭБ, изменения их морфологии при воздействии несептических доз ЛПС маловероятны. Реакция олигодендроцитов показана у новорожденных животных при в/б введении ЛПС 1 мг/кг массы тела [19], а также у животных при прямом стереотаксическом введении ЛПС в структуры головного мозга [20] и при моделировании острого нарушения мозгового кровообращения [21].
Значения представлены в виде медианы и межквартильного размаха (25-й; 75-й процентили). Для межгруппового сравнения использовали U-критерий Манна – Уитни. Уровень статистической значимости в эксперименте принят как p < 0,05. Статистическую обработку данных выполняли при помощи программного обеспечения Microsoft Excel (Microsoft, США) и OriginPro (OriginLab, США).
При гистологическом исследовании срезов головного мозга у мышей обеих групп не выявлено повышенного кровенаполнения сосудов и кровоизлияний. Также ни в одной структуре головного мозга не отмечено периваскулярного отека или миграции лейкоцитов. ИГХ исследование выявило положительную реакцию на антитела к белкам GFAP и Iba1 на срезах коры больших полушарий головного мозга в обеих группах. Экспрессия маркеров GFAP и Iba1 показана на рисунке 2.
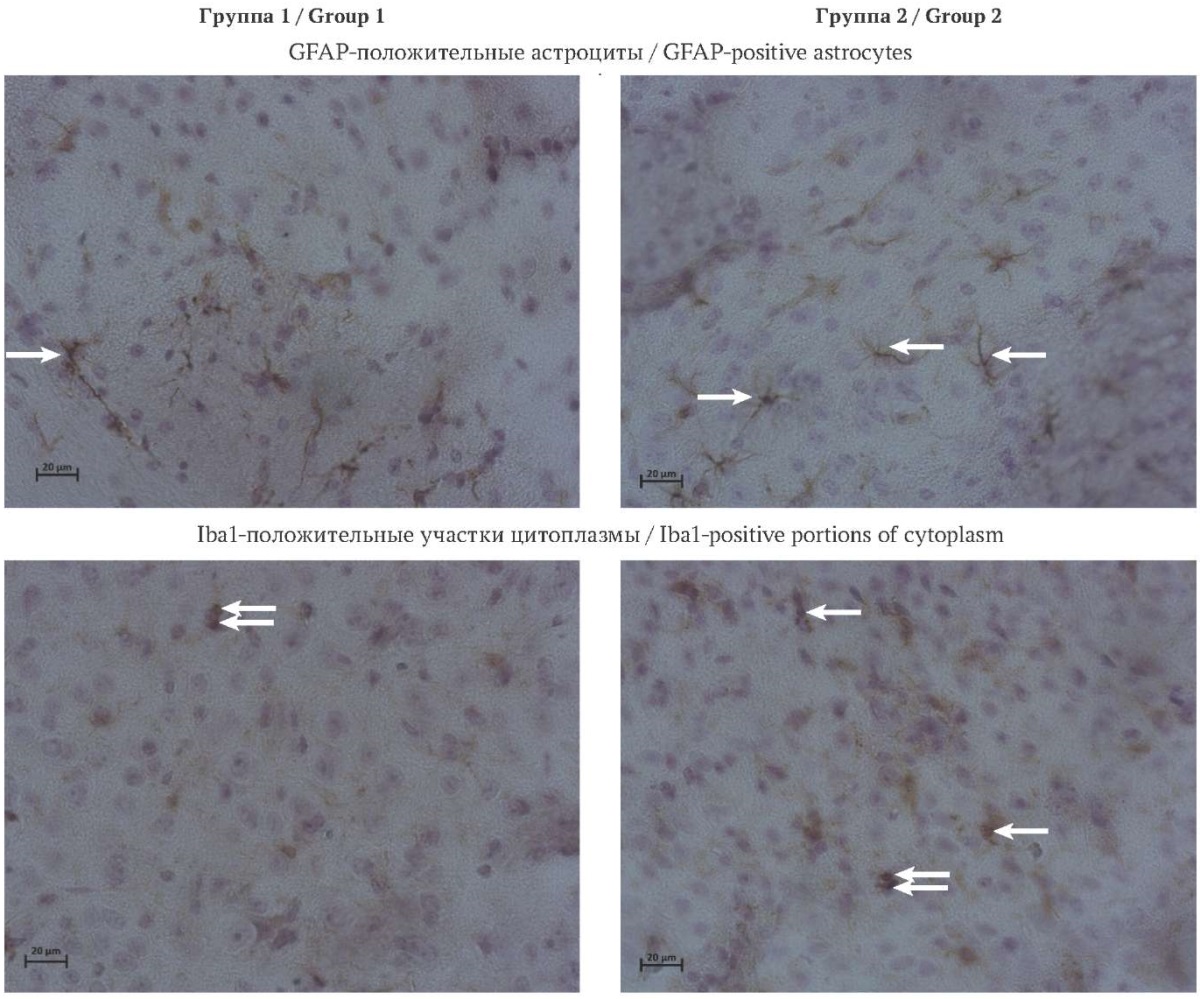
РИС. 2. Гистологические препараты цингулярной и ретросплениальной коры больших полушарий головного мозга мышей на пятые сутки эксперимента. Корональные срезы, d = 5 мкм. Окраска 3,3’-диаминобензидином и гематоксилином Карацци, об.×40.
Примечание: стрелками указаны клетки или участки их цитоплазмы с положительной реакцией антител к GFAP или Iba1, двойными стрелками указаны клетки с несколькими зонами Iba1-положительного окрашивания.
GFAP – glial fibrillary acidic protein, глиальный фибриллярный кислый белок; Iba1 – ionized calcium-binding adapter molecule 1, ионизированная кальций-связывающая адаптерная молекула 1.
FIG. 2. Histological slides of cerebral cingulate and retrosplenial cortex of mice on Day 5 of the experiment. Coronal sections, d = 5 μm. 3,3’-diaminobenzidine and Carazzi hematoxylin staining, ob.×40.
Note: arrows mark cells or portions of their cytoplasm with positive reaction of antibodies to GFAP or Iba1, double arrows mark cells with multiple Iba1-positive areas.
GFAP – glial fibrillary acidic protein; Iba1 – ionized calcium-binding adapter molecule 1.
При подсчете количества клеток, экспрессирующих GFAP, в группе 1 выявлено 9,0 (4,3; 17,0), в группе 2 – 22,5 (8,0; 32,0). Различие между медианными значениями статистически значимо с величиной p = 0,0038 (рис. 3А).
Анализ участков цитоплазмы, экспрессирующих Iba1, выявил 128 (89,3; 165,5) Iba1-положительных участков цитоплазмы в поле зрения в группе 1 и 207,5 (154,8; 295,8) в группе 2 (рис. 3B). Различия между медианными значениями статистически значимы (p = 0,014).
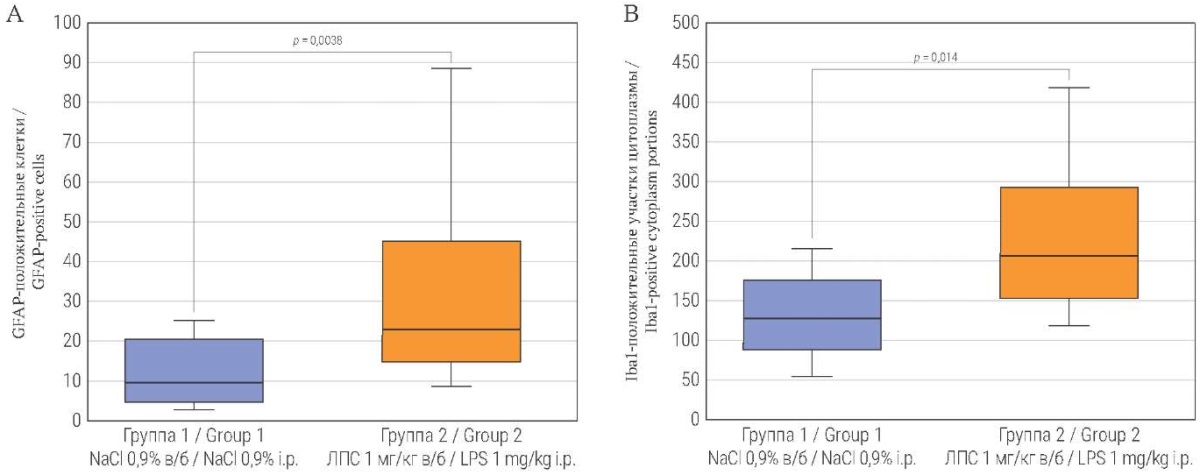
РИС. 3. Количество GFAP-положительных клеток (A) и Iba1-положительных участков цитоплазмы (B) в цингулярной и ретросплениальной коре больших полушарий головного мозга у мышей на пятые сутки эксперимента.
Примечание: GFAP – glial fibrillary acidic protein, глиальный фибриллярный кислый белок; Iba1 – ionized calcium-binding adapter molecule 1, ионизированная кальций-связывающая адаптерная молекула 1; ЛПС – бактериальный липополисахарид; в/б – внутрибрюшинный.
FIG. 3. The number of GFAP-positive cells (A) and the number of Iba1-positive cytoplasm areas (B) in the cingulate and retrosplenial cerebral cortex in mice on Day 5 of the experiment.
Note: GFAP – glial fibrillary acidic protein; Iba1 – ionized calcium-binding adapter molecule 1; LPS – bacterial lipopolysaccharide; i.p. – intraperitoneally.
Полученные результаты свидетельствуют о статистически значимом повышении экспрессии GFAP и Iba1 в цингулярной и ретросплениальной коре больших полушарий головного мозга у мышей после введения ЛПС 1 мг/кг/сутки в течение 4 дней.
Известно, что белок GFAP характерен для промежуточных филаментов в астроцитах [22], хотя его можно выявить не только в астроглии [23]. При гистологическом исследовании в описанном эксперименте увеличение экспрессии GFAP наблюдалось именно в астроцитах мышей экспериментальной группы.
Белок Iba1 характерен для популяций реактивных макрофагов, в том числе и в нейроглии [24]. Кроме того, Iba1 является маркером не только клеток микроглии, но и клеток моноцитарно-макрофагального звена [24][25]. Это обеспечивает широкий спектр применения Iba1, но не позволяет отличить клетки микроглии от макрофагов, если последние проникнут через ГЭБ при воспалительной инфильтрации. В то же время выбранные для эксперимента дозы ЛПС не привели к появлению признаков воспалительной инфильтрации или иных патологических изменений, видимых при световой микроскопии.
Эти данные согласуются с литературными источниками. Так, при режиме введения ЛПС мышам в дозе 2 мг/кг/сутки в/б в течение 3 суток, выбранном для моделирования нарушений аффекта и поведения, происходила активация как астроцитов, так и макрофагов [1]. Аналогичные морфологические изменения отмечены при в/б введении мышам ЛПС 0,25 мг/кг 7 раз в течение двух недель, что сопровождалось развитием повреждения нервной ткани, с помощью которого моделировали нейродегенеративные процессы [26]. Более высокие дозы ЛПС приводят к увеличению экспрессии Iba1 в клетках микроглии уже через несколько часов [27], поскольку ведут к нарушению целостности и проницаемости ГЭБ для большого количества популяций лейкоцитов [28–30].
Результаты проведенного исследования демонстрируют, что воздействие бактериального агента, приводящее к активации нейроглии, доступно количественной оценке по экспрессии ИГХ маркеров. В первые часы после введения ЛПС уровень Iba1 может не изменяться, а уровень GFAP реагирует и восстанавливается быстрее, чем уровень Iba1 [31].
Новизна настоящего исследования состоит в том, что режим введения ЛПС в течение 4 суток показан как эффективный с точки зрения одновременного изучения экспрессии GFAP и Iba1 в клетках нейроглии.
Следует подчеркнуть, что клетки, экспрессирующие GFAP, производят разное количество белков цитоскелета при различных повреждениях [18]. Другими словами, экспрессия GFAP является показателем активации астроцитов при воздействии типовым внешним сигналом повреждения, каковым является ЛПС. Именно такие сигналы в большей степени моделируют возрастные изменения при взаимодействии нейроглии и микробиоты толстой кишки, где E. coli составляет значительную долю бактерий [32].
В последнее время появляются сведения о положительной экспрессии маркера GFAP нейронами в ходе структурных изменений в нервной ткани при травматическом повреждении [33]. В сером веществе коры больших полушарий экспрессия GFAP наступает позже, чем в белом веществе, и в большей степени отражает усиление механической, опорной функции цитоскелета в астроцитах для укрепления ГЭБ [34].
Следует отметить, что введение ЛПС в некоторых случаях сопровождается морфологическими изменениями в нервной ткани головного мозга, например снижением количества клеток, которые могут быть определены при рутинных методах окрашивания (по Нисслю) [35]. Однако увеличение экспрессии ИГХ маркеров GFAP и Iba1 в нервной ткани является более специфичной оценкой.
Размер выборки в эксперименте был ограничен этической целесообразностью и концепцией 3R. Тем не менее полученные данные статистически значимы. Экстраполяция результатов эксперимента, выполненного на мышах, ограниченно возможна на человека.
Структурные изменения в нервной ткани при введении ЛПС могут развиваться в результате воздействия цитокинов, приводящего к изменению в жизненном цикле клеток нейроглии. Представляется перспективным изучение молекулярных каскадов и изменений в популяции нейронов при аналогичных дозе и способе введения ЛПС у мышей, а также реакции нейроглии и нейронов при воздействии других доз, режимах и способах введения ЛПС.
Введение в/б несептической дозы ЛПС в течение 4 суток приводит к астроцитарной и микроглиальной активации: увеличивается количество астроцитов с повышенным содержанием промежуточных филаментов (GFAP-положительных клеток), а также экспрессия маркеров макрофагов (Iba1) в цингулярной и ретросплениальной коре больших полушарий головного мозга. Изученная доза позволяет оценить изменения в ЦНС со стороны глии при повреждении, не сопровождающемся развитием воспалительной реакции.
А.А. Венедиктов, Г.А. Пьявченко и С.Л. Кузнецов сформулировали идею и разработали дизайн исследования. А.А. Венедиктов, Д.М. Оганесян и В.В. Голоборщева изучили литературные источники, провели эксперименты с животными и изготовили гистологические препараты. А.А. Венедиктов выполнил статистическую обработку результатов. А.А. Венедиктов, В.В. Голоборщева, Г.А. Пьявченко и С.Л. Кузнецов написали текст статьи. А.А. Венедиктов и К.С. Покидова оформили иллюстрации. Все авторы утвердили окончательную версию статьи.
Соответствие принципам этики. Исследование проведено с соблюдением положений Европейской конвенции о защите позвоночных животных, которые используются для экспериментальных и других научных целей. Все манипуляции с животными проведены в соответствии с разрешением Локального этического комитета ФГБОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (№ 01-22 от 20.01.2022).
Доступ к данным исследования. Данные, подтверждающие выводы этого исследования, можно получить у авторов по обоснованному запросу.
Данные и статистические методы, представленные в статье, прошли статистическое рецензирование редактором журнала – сертифицированным специалистом по биостатистике.
Конфликт интересов. Авторы заявляют об отсутствии конфликта интересов.
Финансирование. Исследование не имело спонсорской поддержки (собственные ресурсы).
1. Артикул ET1601-23, партия HO0913, Huabio, США. https://huabioresearch.com/datasheet/ET1601-23 (дата обращения: 11.07.2024).
2. Артикул ER1802-20, партия HL0213, Huabio, США. https://huabioresearch.com/datasheet/ER1802-20 (дата обращения: 11.07.2024).
3. Артикул HA1119, партия M05-22-P2, Huabio, США. https://huabioresearch.com/datasheet/HA1119 (дата обращения: 11.07.2024).
4. QuPath Open Software for Bioimage Analysis. https://qupath.github.io (дата обращения 07.10.2024).
1. Li W., Ali T., He K., et al. Ibrutinib alleviates LPS-induced neuroinflammation and synaptic defects in a mouse model of depression. Brain Behav Immun. 2021 Feb; 92: 10–24. https://doi.org/10.1016/j.bbi.2020.11.008. Epub 2020 Nov 10. PMID: 33181270
2. Norden D.M., Trojanowski P.J., Villanueva E., et al. Sequential activation of microglia and astrocyte cytokine expression precedes increased Iba-1 or GFAP immunoreactivity following systemic immune challenge. Glia. 2016 Feb; 64(2): 300–316. https://doi.org/10.1002/glia.22930. Epub 2015 Oct 15. PMID: 26470014
3. Sekio M., Seki K. Lipopolysaccharide-induced depressive-like behavior is associated with α₁-adrenoceptor dependent downregulation of the membrane GluR1 subunit in the mouse medial prefrontal cortex and ventral tegmental area. Int J Neuropsychopharmacol. 2014 Oct 31; 18(1): pyu005. https://doi.org/10.1093/ijnp/pyu005. Erratum in: Int J Neuropsychopharmacol. 2016 Apr 27; 19(10): pyw031. https://doi.org/10.1093/ijnp/pyw031. PMID: 25539502
4. Allen W.E., Blosser T.R., Sullivan Z.A., et al. Molecular and spatial signatures of mouse brain aging at single-cell resolution. Cell. 2023 Jan 5; 186(1): 194–208.e18. https://doi.org/10.1016/j.cell.2022.12.010. Epub 2022 Dec 28. PMID: 36580914.
5. Пьявченко Г.А., Венедиктов А.А., Кузьмин Е.А. и др. Морфофункциональные изменения у мышей после однократного введения высоких и низких доз Hsp70. Сеченовский вестник. 2023. 14(4): 31–41. https://doi.org/10.47093/2218-7332.2023.918.13. Публикация онлайн 29.11.2023. EDN: WALXAM
6. Venediktov A.A., Bushueva O.Y., Kudryavtseva V.A., et al. Closest horizons of Hsp70 engagement to manage neurodegeneration. Front Mol Neurosci. 2023 Sep 19; 16: 1230436. https://doi.org/10.3389/fnmol.2023.1230436. PMID: 37795273
7. Catorce M.N., Gevorkian G. LPS-induced murine neuroinflammation model: main features and suitability for pre-clinical assessment of nutraceuticals. Curr Neuropharmacol. 2016; 14(2), 155–164. https://doi.org/10.2174/1570159x14666151204122017. PMID: 26639457
8. Rolls E.T. The cingulate cortex and limbic systems for emotion, action, and memory. Brain Struct Funct. 2019 Dec; 224(9): 3001–3018. https://doi.org/10.1007/s00429-019-01945-2. Epub 2019 Aug 26. PMID: 31451898
9. Balcerek E., Włodkowska U., Czajkowski R. Retrosplenial cortex in spatial memory: focus on immediate early genes mapping. Mol Brain. 2021 Dec 4; 14(1): 172. https://doi.org/10.1186/s13041-021-00880-w. PMID: 34863215
10. Middeldorp J., Hol E.M. GFAP in health and disease. Progress in neurobiology. 2011; 93(3): 421–443. https://doi.org/10.1016/j.pneurobio.2011.01.005
11. Kierdorf K., Prinz M. Factors regulating microglia activation. Frontiers in cellular neuroscience. 2013; 7: 44. https://doi.org/10.3389/fncel.2013.00044
12. Ohsawa K., Imai Y., Sasaki Y., Kohsaka S. Microglia/macrophage-specific protein Iba1 binds to fimbrin and enhances its actin-bundling activity. Journal of neurochemistry. 2004; 88(4): 844–856. https://doi.org/10.1046/j.1471-4159.2003.02213.x
13. Huang H., He W., Tang T., et al. Immunological markers for central nervous system glia. Neurosci Bull. 2023 Mar; 39(3): 379–392. https://doi.org/10.1007/s12264-022-00938-2. Epub 2022 Aug 26. PMID: 360286412023
14. Balu D., Valencia-Olvera A.C., Nguyen A., et al. A small-molecule TLR4 antagonist reduced neuroinflammation in female E4FAD mice. Alzheimers Res Ther. 2023 Oct 19; 15(1): 181. https://doi.org/10.1186/s13195-023-01330-6. PMID: 37858252
15. Festing M.F.W., Altman D.G. Guidelines for the design and statistical analysis of experiments using laboratory animals. ILAR J. 2002; 43(4): 244–258. https://doi.org/10.1093/ilar.43.4.244. Erratum in: ILAR J. 2005; 46(3): 320. PMID: 12391400
16. Harland M., Torres S., Liu J., et al. Neuronal mitochondria modulation of LPS-induced neuroinflammation. J Neurosci. 2020 Feb 19; 40(8): 1756–1765. https://doi.org/10.1523/JNEUROSCI.2324-19.2020. Epub 2020 Jan 14. PMID: 31937559
17. Paxinos G., Franklin K.B.J. The Mouse Brain in Stereotaxic Coordinates. 5th Edition. Academic Press. 2019. 376p. eBook ISBN: 9780128161586
18. Bankhead P., Loughrey M.B., Fernández J.A., et al. QuPath: Open-source software for digital pathology image analysis. Sci Rep. 2017 Dec 4; 7(1): 16878. https://doi.org/10.1038/s41598-017-17204-5. PMID: 29203879
19. Huang P., Zhou Q., Lin Q., et al. Complement C3a induces axonal hypomyelination in the periventricular white matter through activation of WNT/β-catenin signal pathway in septic neonatal rats experimentally induced by lipopolysaccharide. Brain Pathol. 2020 May; 30(3): 495–514. https://doi.org/10.1111/bpa.12798. Epub 2019 Nov 6. PMID: 31622511
20. Yao S.Y., Ljunggren-Rose A., Chandramohan N., et al. In vitro and in vivo induction and activation of nNOS by LPS in oligodendrocytes. Journal of Neuroimmunology. 2010; 229(1-2): 146–156. https://doi.org/10.1016/j.jneuroim.2010.07.023
21. Cammarota M., Boscia F. Contribution of oligodendrocytes, microglia, and astrocytes to myelin debris uptake in an explant model of inflammatory demyelination in rats. Cells. 2023 Sep 3; 12(17): 2203. https://doi.org/10.3390/cells12172203. PMID: 37681935
22. Romero-Sandoval A., Chai N., Nutile-McMenemy N., Deleo J.A. A comparison of spinal Iba1 and GFAP expression in rodent models of acute and chronic pain. Brain research. 2008; 1219: 116–126. https://doi.org/10.1016/j.brainres.2008.05.004
23. Oberheim N.A., Goldman S.A., Nedergaard M. Heterogeneity of astrocytic form and function. Methods Mol Biol. 2012; 814: 23–45. https://doi.org/10.1007/978-1-61779-452-0_3. PMID: 2214429
24. Woodburn S.C., Bollinger J.L., Wohleb E.S. The semantics of microglia activation: neuroinflammation, homeostasis, and stress. J Neuroinflammation. 2021 Nov 6; 18(1): 258. https://doi.org/10.1186/s12974-021-02309-6. PMID: 34742308
25. Zhang X., Wang L.P., Ziober A., et al. Ionized Calcium Binding Adaptor Molecule 1 (IBA1). Am J Clin Pathol. 2021 Jun 17; 156(1): 86–99. https://doi.org/10.1093/ajcp/aqaa209. PMID: 33582751
26. Muhammad T., Ikram M., Ullah R., et al. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients. 2019 Mar 17; 11(3): 648. https://doi.org/10.3390/nu11030648. PMID: 30884890
27. Arioz B.I., Tastan B., Tarakcioglu E., et al. Melatonin attenuates LPS-induced acute depressive-like behaviors and microglial NLRP3 inflammasome activation through the SIRT1/Nrf2 pathway. Front Immunol. 2019 Jul 2; 10: 1511. https://doi.org/10.3389/fimmu.2019.01511. PMID: 31327964
28. Dumbuya J.S., Chen X., Du J., et al. Hydrogen-rich saline regulates NLRP3 inflammasome activation in sepsis-associated encephalopathy rat model. Int Immunopharmacol. 2023 Oct; 123: 110758. https://doi.org/10.1016/j.intimp.2023.110758; Epub 2023 Aug 8. PMID: 37556997
29. Park J.H., Hwang J.W., Lee H.J., et al. Lomerizine inhibits LPS-mediated neuroinflammation and tau hyperphosphorylation by modulating NLRP3, DYRK1A, and GSK3α/β. Front Immunol. 2023 Jun 26; 14: 1150940. https://doi.org/10.3389/fimmu.2023.1150940. PMID: 37435081
30. Xingi E., Koutsoudaki P.N., Thanou I., et al. LPS-induced systemic inflammation affects the dynamic interactions of astrocytes and microglia with the vasculature of the mouse brain cortex. Cells. 2023 May 17; 12(10): 1418. https://doi.org/10.3390/cells12101418. PMID: 374082522023
31. Anderson M.A., Ao Y., Sofroniev M.V. Heterogeneity of reactive astrocytes. Neuroscience letters. 2014; 565: 23–29. https://doi.org/10.1016/j.neulet.2013.12.030
32. Mayer E.A., Nance K., Chen S. The gut-brain axis. Annu Rev Med. 2022 Jan 27; 73: 439–453. https://doi.org/10.1146/annurev-med-042320-014032. Epub 2021 Oct 20. PMID: 34669432022
33. Zwirner J., Lier J., Franke H., et al. GFAP positivity in neurons following traumatic brain injuries. Int J Legal Med. 2021 Nov; 135(6): 2323–2333. https://doi.org/10.1007/s00414-021-02568-1. Epub 2021 Jun 11. PMID: 34114049
34. Jurga A.M., Paleczna M., Kadluczka J., et al. Beyond the GFAP-astrocyte protein markers in the brain. Biomolecules. 2021 Sep 14; 11(9): 1361. https://doi.org/10.3390/biom11091361. PMID: 345725722021
35. Weng J., Wang Y., Tan Z., et al. Glabridin reduces neuroinflammation by modulating inflammatory signals in LPS-induced in vitro and in vivo models. Inflammopharmacology. 2024 Apr; 32(2): 1159–1169. https://doi.org/10.1007/s10787-023-01424-5. Epub 2024 Feb 19. PMID: 38372849
Венедиктов Артем Андреевич - аспирант, ассистент кафедры анатомии и гистологии человека ФГАОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (Сеченовский Университет).
ул. Трубецкая, д. 8, стр. 2, Москва, 119048
Покидова Ксения Сергеевна - студентка ФГАОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (Сеченовский Университет).
ул. Трубецкая, д. 8, стр. 2, Москва, 119048
Оганесян Давид Михайлович - студент ФГАОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (Сеченовский Университет).
ул. Трубецкая, д. 8, стр. 2, Москва, 119048
Голоборщева Валерия Владимировна - канд. мед. наук, младший научный сотрудник лаборатории общей патологии нервной системы ФГБНУ «НИИОПП».
ул. Балтийская, д. 8, Москва, 125315
Пьявченко Геннадий Александрович - канд. мед. наук, доцент кафедры анатомии и гистологии человека ФГАОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (Сеченовский Университет).
ул. Трубецкая, д. 8, стр. 2, Москва, 119048
Кузнецов Сергей Львович - член-корр. РАН, д-р мед. наук, профессор кафедры анатомии и гистологии человека ФГАОУ ВО «Первый МГМУ им. И.М. Сеченова» Минздрава России (Сеченовский Университет).
ул. Трубецкая, д. 8, стр. 2, Москва, 119048

|
1. The ARRIVE guidelines 2.0: author checklist | |
| Тема | ||
| Тип | Исследовательские инструменты | |
Скачать
(136KB)
|
Метаданные ▾ | |
119048, г. Москва, ул. Трубецкая, д. 8, стр. 2
E-mail: sechenovmedj@staff.sechenov.ru

































