




Содержание
Перейти к:
https://doi.org/10.47093/2218-7332.2023.14.3.28-36
Перейти к:
Цель. Определить содержание нейронов, экспрессирующих соматостатин (SST), и его колокализацию с тирозингидроксилазой (TH) и нейропептидом Y (NPY) в краниальном шейном ганглии (КШГ) и солнечном сплетении крысы.
Материалы и методы. Работа выполнена на 30 белых крысах-самцах линии Wistar шести возрастных групп (по 5 крыс в каждой): новорожденные, 10-, 20-, 30-, 60-суточные, 24-месячные. Срезы ганглиев инкубировали с первичными антителами к SST, NPY, TH и вторичными, конъюгированными с флуорохромами. Определяли долю иммунореактивных (IR) нейронов с видимым ядрышком и с флуоресценцией, превышающей фоновое свечение, по отношению к общему числу нейронов, а также среднюю площадь сечения с помощью программы Image J (NIH, США).
Результаты. SST-IR нейроны отсутствовали в КШГ. Иммунореактивный материал обнаруживался в области тел большинства нейронов солнечного сплетения в виде гранул для SST и NPY и располагался относительно гомогенно для TH. Доля SST-IR нейронов составляла 33% у новорожденных, увеличивалась в два раза в первый месяц жизни и далее оставалась постоянной (70–73%). Не установлено статистически значимых различий по долям SST-IR нейронов между краниальным брыжеечным ганглием (КБГ) и чревным ганглием (ЧГ) во всех возрастных группах. С момента рождения и до 60 суток жизни средняя площадь сечения SST-IR нейронов в ЧГ и КБГ увеличивалась в 3,4–3,9 раза и далее не менялась до 24 мес. С 20-х суток жизни средняя площадь сечения SST-IR нейронов в ЧГ была статистически значимо выше, чем в КБГ. Все SST-IR нейроны во всех возрастных группах содержали TH и 90–94% нейронов колокализовали NPY.
Заключение. Содержание SST-IR нейронов в различных симпатических узлах гетерохронно: они отсутствуют в КШГ, а их доля и площадь в солнечном сплетении в раннем постнатальном онтогенезе увеличивается. Это может быть связано с особенностями иннервируемых органов-мишеней
Соматостатин (somatostatin, SST) представляет собой полипептид, относящийся к гормонам и нейропептидам. Существует в двух биологически активных формах: SST-14 и SST-28, которые различаются, соответственно, числом входящих в состав молекулы аминокислот [1]. Физиологические функции опосредованы 5 типами рецепторов к соматостатину: SSTR1–SSTR5 (somatostatin receptor), которые имеют похожую структуру и механизмы передачи сигналов, но отличаются своей клеточной и субклеточной локализацией и способом регуляции [1][2].
SST прежде всего рассматривается как сильный ингибирующий фактор в эндокринной системе и желудочно-кишечном тракте; влияет на деление клеток, сократимость гладких мышц внутренних органов, обмен питательными веществами. В качестве нейропептида SST оказывает прямое действие на центральную нервную систему и передачу нервных импульсов [1][3]; также играет роль трофического фактора в развитии нейронов в эмбриогенезе [4]. В недавних исследованиях обнаружено присутствие SST в автономной нервной системе млекопитающих – в части нейронов симпатических узлов [5]. В подавляющем большинстве этих нейронов содержится медиатор норадреналин и фермент его синтеза тирозингидроксилаза (tyrosine hydroxylase, TH), две трети нейронов содержат нейропептид Y (neuropeptide Y, NPY) [6][7].
Нейромедиаторный состав симпатических нейронов изменяется в онтогенезе, а также различен по локализации в ганглиях [8–10]. В частности, у крысы в эмбриогенезе на 16 сутки эмбрионального развития наблюдается транзиторная экспрессия SST в большинстве симпатических ганглионарных нейронов [10][11].
Во время эмбрионального развития морской свинки в солнечном сплетении SST-иммунореактивные (immunoreactive, IR) нейроны появляются позже, чем NPY-IR, причем большинство (75%) SST-IR нейронов на поздних эмбриональных стадиях продолжают экспрессировать NPY, что в неонатальном периоде наблюдается редко (<2%) [8][12].
После рождения у мыши в превертебральных узлах выявляется малое число SST-IR нейронов [13]. тогда как у морской свинки их количество в превертебральных узлах солнечного сплетения достигает 25%, а у свиней 12–15%, в отличие от паравертебральных узлов [14][15]. Так, в паравертебральном звездчатом ганглии крысы самая высокая доля нейронов, содержащих SST, наблюдается у новорожденных (7% от общего числа нейронов), позже количество SST-IR нейронов снижается до 4% и остается постоянным, начиная с 10 дней жизни [9].
В литературных источниках отсутствуют данные о постнатальных изменениях состава нейромедиаторов в других симпатических узлах, в частности паравертебральном краниальном шейном ганглии (КШГ), а также в солнечном сплетении: превертебральных чревных ганглиях (ЧГ) и краниальном брыжеечном ганглии (КБГ).
Целью исследования является определение содержания SST-IR нейронов в КШГ и солнечном сплетении крысы в постнатальном онтогенезе, а также колокализации соматостатина с другими нейротрансмиттерами, включая фермент синтеза катехоламинов TH и NPY.
Работа выполнена на 30 белых крысах-самцах линии Wistar следующих шести возрастных групп (по 5 крыс в каждой): новорожденные, 10-, 20-, 30-, 60-суточные, 24-месячные.
Животных выводили из эксперимента путем инъекции летальной дозы уретана (3 г/кг, внутрибрюшинно). Далее проводилась транскардиальная перфузия раствором стандартного фоcфатно-cолевого буфеpа (phosphate buffered saline, PBS) 0,01 М, pH 7,4 (ООО «БиолоТ», РФ) объемом от 20 до 500 мл в зависимости от возраста, а затем таким же объемом фиксирующей смеси 4% pаcтвоpа паpафоpмальдегида (Sigma, США) в PBS. Далее извлекались: КШГ, ганглии солнечного сплетения (правый и левый ЧГ и КБГ) и помещались на 2 часа при комнатной температуре в ту же фиксирующую смесь, в которой проводили перфузию. Затем ганглии три раза промывались в PBS по 10 минут с интервалом в 5 минут и перемещались в 30% раствор сахарозы на PBS на 12 часов при температуре 4 °С. Серии срезов толщиной 12 мкм изготовляли на криостате.
На следующем этапе проводили прединкубацию срезов в течение 30 мин при комнатной температуре в PBS с добавлением 10% ослиной сыворотки (Jackson Immunoresearch, США), 1% тритона X-100, 0,1% бычьего сывороточного альбумина, 0,05% тимерозола. После этого срезы инкубировали с первичными антителами: козьими против SST, 1:200 (Santa Cruz, sc-7819), овечьими против TH, 1:1000 (Abcam, ab113), кроличьими против NPY, 1:500 (Abcam, ab30914), в течение 24 ч при комнатной температуре. После кратковременной промывки в PBS срезы инкубировали со вторичными антителами в течение 2 ч. Вторичные антитела были конъюгированы с флуорохромами – флуоресцеин изотиоцианатом (fluorescein isothiocyanate, FITC), дающим зеленую флуоресценцию, и индокарбоцианином (indocarbocyanine, CY3), дающим красную флуоресценцию (разведение 1:150, Jackson Immunoresearch, США).
Окраску клеток всей популяции нейронов проводили зеленым флуоресцентным красителем NeuroTrace™ Green Fluorescent Nissl Stains (Molecular Probes, США) – разведение 1:200. После этого срезы отмывали в PBS и заключали в среду для флуоресцентной микроскопии VectaShield (Vector Laboratories, США). Для исключения неспецифической реакции часть срезов инкубировали без первичных и/или вторичных антител.
Анализ препаратов проводился с использованием программно-аппаратного комплекса, включающего флуоресцентный микроскоп Olympus BX43 (Olympus, Япония) с набором фильтров (модуль зеркал UFBWA – синее возбуждение, фильтр возбуждения BP460-495, барьерный фильтр BA510550 – зеленая флуоресценция; модуль зеркал U-FGWА – зеленое возбуждение, фильтр возбуждения BP530-550, барьерный фильтр BA575-625 – красная флуоресценция), охлаждаемую ССD камеру Tucsen FL-20 (Xintu Photonics, Китай) и компьютер на базе процессора Intel Core i7 (Intel, США). Получение и последующая обработка изображений проводилась с использованием программы Mosaic, версия 2.1 (Xintu Photonics, Китай).
Полученные мультиканальной флуоресценцией с различными спектрами флуорохромов изображения препаратов комбинировали при помощи программы Paint Shop Pro 7.02 (Jasc Software, Inc., США), при этом накладывали снимки одного и того же поля зрения. Объединенный рисунок отображал зеленый и красный цвета и зоны их перекрытия в виде градиента желто-зеленого цвета.
Для выявления меченых нейронов использовали каждый третий из серийных срезов. Число меченых нейронов определяли на изображениях срезов площадью 0,14 мм², полученных под объективом 20х/0,50. Долю SST-IR нейронов определяли как их отношение к общему количеству нейронов, выявленных с помощью NeuroTrace™ Fluorescent NisslStains, которое принимали за 100%. Анализу подлежали нейроны, срез которых прошел через ядро с видимым ядрышком и с флуоресценцией, превышающей фоновое свечение. Число меченых нейронов, а также среднюю площадь сечения SST-IR нейронов определяли с помощью программы Image J (NIH, США). Для определения средней площади сечения при увеличении 200 вручную обводили границы IR клеток. Всего для анализа брали 200 нейронов в каждом ганглии каждой возрастной группы.
Проверка на нормальность распределения случайных величин проводилась с использованием критерия Шапиро – Уилка. Значимость различий средних величин оценивалась при помощи однофакторного дисперсионного анализа (ANOVA). Для парных сравнений использовался t-критерий Стьюдента, для множественных межгрупповых сравнений использовалась поправка Бонферрони. Различия принимались как значимые при р < 0,05. Для статистического анализа данных использовалась программа Sigma Plot (Systat Software, США).
Результаты показали, что иммуномечение к SST и NPY было представлено в виде гранул, заполняющих цитоплазму, в то время как иммунореактивность к TH была относительно гомогенной. В большинстве нейронов иммунореактивный материал обнаруживался в области тел, тем не менее у отдельных клеток он выявлялся и в области проксимальных дендритов.
Меченые нейроны были распределены по поверхности среза относительно равномерно. Мы не обнаружили количественных различий между левым и правым ЧГ, поэтому далее представлены усредненные данные по обоим узлам.
SST-IR нейроны не обнаруживались в КШГ, но выявлялись в солнечном сплетении. Начиная с периода новорожденности и до 24 мес. часть нейронов и в ЧГ, и в КБГ была SST-IR (рис. 1).
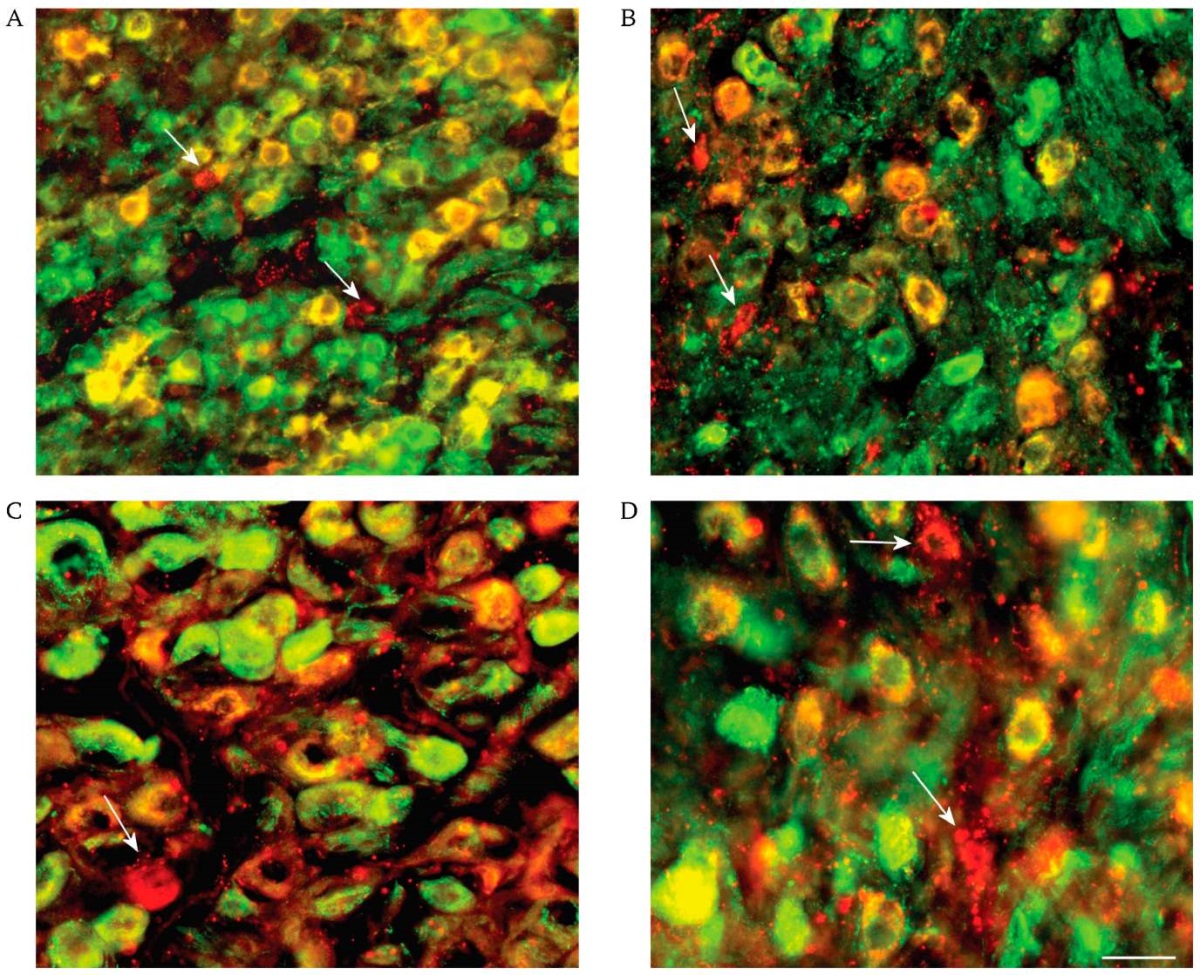
РИС. 1. Содержание в чревном ганглии у крыс разных возрастов нейронов,
экспрессирующих соматостатин (красные) и нейропептид Y (зеленые),
и их колокализация (желтый цвет).
Микрофото: об. 20, ок. 10. Масштаб: 30 мкм.
Иммунонегативные к нейропептиду Y
соматостатинергические нейроны указаны стрелкой.
Флуоресценция: флуоресцеин-изотиоцианат и индокарбоцианин.
Возраст крысы: A – новорожденная, B – 10-суточная,
C – 60-суточная, D – 24-месячная.
FIG. 1. Content of neurons expressing somatostatin (red)
and neuropeptide Y (green) and their colocalization (yellow)
in celiac ganglia of rats at different ages.
Microphotograph: ob. 20, oc. 10. Scale: 30 µm.
Somatostatinergic neurons immunonegative for neuropeptide Y
are indicated by arrows.
Fluorescence: fluorescein isothiocyanate and indocarbocyanine.
Age of the rat: A – newborn pup, B – 10-day-old,
C – 60-day-old, D – 24-month-old.
У новорожденных треть нейронов в ЧГ и КБГ была с IR. Доля SST-IR нейронов удваивалась в онтогенезе в обоих узлах в течение первых 30 суток жизни и сохранялась на уровне 70–73% до 24 мес. (табл. 1). Мы не наблюдали значимых различий по долям SST-IR между КБГ и ЧГ в каждой из возрастных групп.
Таблица 1. Доля соматостатинергических нейронов
в симпатических узлах крыс в постнатальном онтогенезе
Table 1. The content of somatostatinergic neurons
in postnatal sympathetic ganglia of rats
|
Возраст / Age |
Доля SST-IR нейронов, % / The content of SST-IR neurons, % |
Значение р / p value |
|
|
Чревный ганглий / Celiac ganglion |
Краниальный брыжеечный ганглий / Cranial mesenteric ganglion |
||
|
Новорожденный / Newborn pup |
34 ± 3,1a |
33 ± 3,3 a |
n.s. |
|
10 суток / 10-day-old |
42 ± 2,7 a |
45 ± 4,1a |
n.s. |
|
20 суток / 20-day-old |
55 ± 4,9 a |
53 ± 5,8 a |
n.s. |
|
30 суток / 30-day-old |
70 ± 6,2 |
72 ± 7,9 |
n.s. |
|
60 суток / 60-day-old |
73 ± 5,9 |
73 ± 6,2 |
n.s. |
|
24 месяца / 24-month-old |
71 ± 5,6 |
71 ± 7,1 |
n.s. |
Примечание: доля соматостатинергических нейронов
представлена в процентах от общего числа нейронов.
Данные по чревному ганглию представлены усредненными значениями
по правому и левому узлам.
a p < 0,05 по сравнению с 60-суточным.
SST-IR – somatostatin immunoreactive, соматостатин иммунореактивные;
n.s. – not significant (не значимо).
Note: the content of somatostatinergic neurons is given
as a percentage of the total number of neurons.
Data for celiac ganglia are presented as average values
for right and left nodes.
a p < 0,05 compared to 60-day-old.
SST-IR – somatostatin immunoreactive; n.s. – not significant.
Средняя площадь сечения SST-IR нейронов увеличивалась в ЧГ и КБГ с момента рождения до 60 суток жизни (табл. 2). С 20-х суток жизни у крыс средняя площадь сечения SST-IR нейронов в ЧГ была статистически значимо выше, чем в КБГ.
Таблица 2. Площадь поперечного сечения
соматостатинергических нейронов в симпатических узлах
крыс разных возрастов
Table 2. Cross-sectional area of somatostatinergic neurons
in the sympathetic ganglions of rats at different ages
|
Возраст / Age |
Площадь поперечного сечения SST-IR нейронов, мкм² / |
Значение р / p value |
|
|
Чревный ганглий / Celiac ganglion |
Краниальный брыжеечный ганглий / Cranial mesenteric ganglion |
||
|
Новорожденный / Newborn pup |
172 ± 7,2a |
167 ± 8,1a |
n.s. |
|
10 суток / 10-day-old |
272 ± 10,7a |
273 ± 12,6a |
n.s. |
|
20 суток / 20-day-old |
453 ± 11,2a |
382 ± 15,5a |
<0,05 |
|
30 суток / 30-day-old |
574 ± 22,2a |
485 ± 19,8a |
<0,05 |
|
60 суток / 60-day-old |
673 ± 33,3 |
576 ± 21,4 |
<0,05 |
|
24 месяца / 24-month-old |
735 ± 34,3 |
622 ± 38,5 |
<0,05 |
Примечание: представлена средняя площадь 200 нейронов.
Данные по чревному ганглию представлены усредненными значениями
по правому и левому узлам.
a p < 0,05 по сравнению с 60-суточным.
SST-IR – somatostatin immunoreactive, соматостатин иммунореактивные;
n.s. – not significant (не значимо).
Note: represents the average area of 200 neurons.
Celiac ganglion data are presented as average values for the right and left nodes.
a p < 0.05 compared to 60-day-old.
SST-IR – somatostatin immunoreactive; n.s. – not significant.
Во всех возрастных группах, с момента рождения и до 24 мес., все SST-IR нейроны ЧГ и КБГ колокализовали фермент синтеза катехоламинов TH. При этом, независимо от возраста, подавляющее большинство SST-IR нейронов (90–94%) содержали NPY (рис. 1, 2). Не было обнаружено значимых различий между долями SST-IR нейронов, колокализующих NPY, в различных возрастных группах (рис. 2).
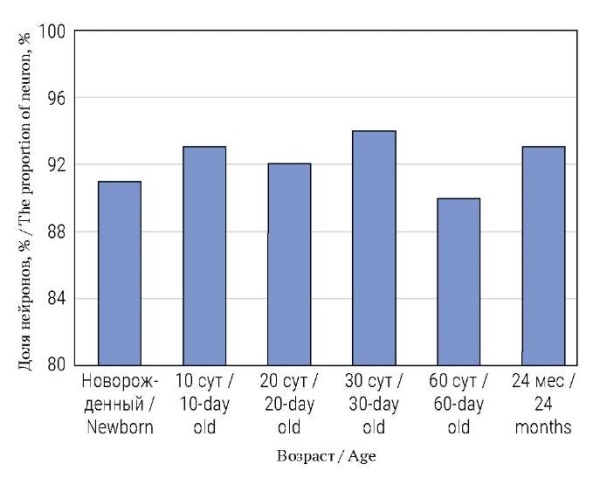
РИС. 2. Процент соматостатинергических (SST) нейронов
в чревном ганглии в постнатальном онтогенезе,
колокализующих нейропептид Y (NPY).
FIG. 2. Percentage of somatostatinergic (SST) neurons
of the celiac ganglion colocalizing neuropeptide Y (NPY)
in postnatal ontogenesis.
Результаты настоящего исследования демонстрируют наличие SST-IR нейронов в превертебральных ЧГ и КБГ к моменту рождения, при этом в паравертебральном КШГ SST-IR нейроны не обнаружены. Доля SST-IR нейронов в превертебральных узлах у крысы превосходит аналогичный показатель у других млекопитающих, включая мышь, морскую свинку, свинью и человека: у мыши и человека обнаруживались лишь отдельные SST-IR нейроны, а у свиней и морской свинки количество SST-IR нейронов в превертебральных узлах доходит до 12–15 и 25% соответственно [5][6][13][14].
Наши более ранние данные свидетельствуют о том, что у крысы в узлах солнечного сплетения содержится больше SST-IR нейронов по сравнению с другими узлами, включая звездчатый узел [9]. Мы впервые обнаружили, что в онтогенезе крысы в первые 30 суток жизни доля SST-IR нейронов увеличивается в превертебральных симпатических узлах, в частности ЧГ и КБГ. В отличие от этого в паравертебральном звездчатом ганглии крыс наибольшее количество нейронов, содержащих SST, обнаруживалось у новорожденных животных и составляло 7% [9] В постнатальном онтогенезе в данном узле доля SST-IR нейронов уменьшается в первые 10 суток жизни до 4% [9]. Таким образом, в различных симпатических узлах доля SST-IR нейронов изменяется гетерохронно, что может быть связано с особенностями иннервируемых органов-мишеней. КШГ обеспечивает симпатическую иннервацию головы, звездчатый ганглий – органов грудной полости и шеи, а превертебральные узлы – органов брюшной полости [16][17]. Ранее нами было показано, что SST-IR нейроны, иннервирующие сердце, выявляются в звездчатом узле только у новорожденных крыс и не наблюдаются у более взрослых [18].
В данной работе мы установили, что доля SST-IR нейронов в превертебральных симпатических узлах крысы начиная с возраста 30 суток далее не изменяется до 2 лет. Согласно литературным данным, в гипоталамусе доля SST-IR нейронов у возрастных особей (2–2,5 года) этого вида животных также оставалась постоянной [19].
Продемонстрированная в данной работе более высокая средняя площадь сечения SST-IR нейронов в ЧГ по сравнению с КБГ у 20-суточных и более взрослых крыс подтверждает обнаруженные ранее данные о больших размерах NPY-IR нейронов в ЧГ относительно КБГ [8]. Нами установлено, что практически все SST-IR нейроны превертебральных узлов крыс в различные возрастные периоды также содержат TH и NPY. Нейромедиаторы SST и NPY подавляют перистальтику и секрецию в желудочно-кишечном тракте [7]. Предполагается, что NPY-IR нейроны солнечного сплетения, не содержащие SST, являются вазомоторными, а NPY/SST-IR – висцеромоторными [20]. Наибольшее число висцеромоторных волокон направляется к интрамуральным узлам межмышечного сплетения желудочно-кишечного тракта [7].
Экспрессия SST может изменяться при патологии – например, у свиньи при воспалении желудочно-кишечного тракта, включая язвенный колит и химически индуцированное воспаление нисходящей ободочной кишки, увеличивается количество данного пептида в ганглионарных нейронах [21]. Имеются данные о том, что SST подавляет воспалительные реакции в кишечнике путем двунаправленной коммуникации между нейронами и тучными клетками, также модулирует активность лимфоидной ткани, ассоциированной с кишечником [22]. SST стимулирует пролиферацию B-лимфобластов, при этом ингибирует активность Т-лимфоцитов, пролиферацию гранулоцитов, синтез провоспалительных цитокинов [23]. SST также участвует в соматостатинергической противовоспалительной петле (somatostatinergic anti-inflammatory loop) [24]: когда повреждающие агенты раздражают пептидергические нейроны, это вызывает высвобождение вещества P и/или кальцитонин-ген-родственного пептида (calcitonin gene-related peptide, CGRP). Последние индуцируют локальное нейрогенное воспаление, которое, в свою очередь, вызывает выброс SST из тех же нейронов. SST замыкает круг, останавливая высвобождение вышеуказанных нейропептидов, и ингибирует воспалительные процессы [24].
В раннем постнатальном онтогенезе крысы в превертебральных узлах наблюдается увеличение доли нейронов, содержащих SST, что важно для становления симпатической иннервации желудочно-кишечного тракта. SST не только тормозит моторику и секрецию, но и оказывает обезболивающее и противовоспалительное действие в органах пищеварения. Дальнейшие эксперименты с изучением в различные возрастные периоды состава SST-рецепторов в органах-мишенях позволят уточнить особенности симпатической регуляции пищеварительной системы и разработать возможные пути фармакологической коррекции заболеваний желудочно-кишечного тракта.
А.И. Емануйлов и В.В. Порсева участвовали в сборе материала, проведении экспериментов и статистической обработке данных. А.В. Павлов и П.М. Маслюков разработали идею и дизайн исследования, а также участвовали в написании рукописи. Все авторы утвердили окончательную версию публикации.
Соответствие принципам этики. Исследование проведено с соблюдением положений Европейской конвенции о защите позвоночных животных, которые используются для экспериментальных и других научных целей. Все манипуляции с животными проведены в соответствии с разрешением Локального этического комитета ФГБОУ ВО «Ярославский государственный медицинский университет» Минздрава России (№ 58-16.05.2022).
Доступ к данным исследования. Данные, подтверждающие выводы этого исследования, можно получить у авторов по обоснованному запросу.
Данные и статистические методы, представленные в статье, прошли статистическое рецензирование редактором журнала – сертифицированным специалистом по биостатистике.
Конфликт интересов. Авторы заявляют об отсутствии конфликта интересов.
Финансирование. Работа выполнена при поддержке Российского научного фонда (РНФ), грант 23-25-00141.
1. Kumar U., Singh S. Role of somatostatin in the regulation of central and peripheral factors of satiety and obesity. Int J Mol Sci. 2020; 21(7): 2568. https://doi.org/10.3390/ijms21072568
2. Günther T., Tulipano G., Dournaud P., et al. International union of basic and clinical pharmacology. CV. Somatostatin receptors: structure, function, ligands, and new nomenclature. Pharmacol Rev. 2018; 70: 763–835. https://doi.org/10.1124/pr.117.015388
3. Ampofo E., Nalbach L., Menger M.D., Laschke M.W. Regulatory mechanisms of somatostatin expression. Int J Mol Sci. 2020; 21(11): 4170. https://doi.org/10.3390/ijms21114170
4. Singh S., Somvanshi R.K., Kumar U. Somatostatin-mediated regulation of retinoic acid-induced differentiation of SHSY5Y cells: neurotransmitters phenotype characterization. Biomedicines. 2022; 10(2): 337. https://doi.org/10.3390/biomedicines10020337
5. Ernsberger U., Deller T., Rohrer H. The diversity of neuronal phenotypes in rodent and human autonomic ganglia. Cell Tissue Res. 2020; 382(2): 201–231. https://doi.org/10.1007/s00441-020-03279-6
6. Palus K., Bulc M., Czajkowska M., et al. Neurochemical characteristics of calbindin-like immunoreactive coeliac-cranial mesenteric ganglion complex (CCMG) neurons supplying the pre-pyloric region of the porcine stomach. Tissue Cell. 2018; 50: 8–14. https://doi.org/10.1016/j.tice.2017.12.002
7. Furness J.B., Di Natale M., Hunne B., et al. The identification of neuronal control pathways supplying effector tissues in the stomach. Cell Tissue Res. 2020; 382(3): 433–445. https://doi.org/10.1007/s00441-020-03294-7
8. Masliukov P.M., Emanuilov A.I., Budnik A.F. Sympathetic innervation of the development, maturity, and aging of the gastrointestinal tract. Anat Rec (Hoboken). 2022; 306 (9): 2249–2263. https://doi.org/10.1002/ar.25015
9. Маслюков П.М., Емануйлов А.И., Ноздрачёв А.Д. Возрастные изменения нейротрансмиттерного состава нейронов симпатических узлов. Успехи геронтологии. 2016; 29(3): 442–453. PMID: 28525692
10. Ernsberger U., Deller T., Rohrer H. The sympathies of the body: functional organization and neuronal differentiation in the peripheral sympathetic nervous system. Cell Tissue Res. 2021. 386(3): 455–475. https://doi.org/10.1007/s00441-021-03548-y
11. Huang T., Hu J., Wang B., et al. Tlx3 controls cholinergic transmitter and Peptide phenotypes in a subset of prenatal sympathetic neurons. J Neurosci. 2013; 33(26): 10667–10675. https://doi.org/10.1523/JNEUROSCI.0192-13.2013. PMID: 23804090
12. Anderson R.L., Morris J.L., Gibbins I.L. Neurochemical differentiation of functionally distinct populations of autonomic neurons. J Comp Neurol. 2001. 429(3): 419–435. https://doi.org/10.1002/1096-9861(20010115)429:3<419::aidcne5>3.0.co;2-d. PMID: 11116229
13. Kaestner C.L., Smith E.H., Peirce S.G., Hoover D.B. Immunohistochemical analysis of the mouse celiac ganglion: An integrative relay station of the peripheral nervous system. J Comp Neurol. 2019; 527(16): 2742–2760. https://doi.org/10.1002/cne.24705
14. Szurszewski J.H., Linden D.R. Physiology of prevertebral sympathetic ganglia in: physiology of the gastrointestinal tract (Johnson L.R., ed). San Diego: Academic Press, 2012. P 583– 627. ISBN 9780123820266. https://doi.org/10.1016/B978-0-12-382026-6.00020-8
15. Furness J.B. Integrated neural and endocrine control of gastrointestinal function. Adv Exp Med Biol. 2016; 891: 159–173. https://doi.org/10.1007/978-3-319-27592-5_16
16. Martinez-Sanchez N., Sweeney O., Sidarta-Oliveira D., et al. The sympathetic nervous system in the 21st century: Neuroimmune interactions in metabolic homeostasis and obesity. Neuron. 2022; 110(21): 3597–3626. https://doi.org/10.1016/j.neuron.2022.10.017
17. Li Y.L. Stellate Ganglia and cardiac sympathetic overactivation in heart failure. Int J Mol Sci. 2022; 23(21): 13311. https://doi.org/10.3390/ijms232113311
18. Емануйлов А.И. Маслюков П.М., Ноздрачев А.Д. Симпатическая иннервация сердца в раннем постнатальном онтогенезе. Российский физиологический журнал им. И.М. Сеченова. 2019; 105(9): 1133–1141. https://doi.org/10.1134/S086981391909005X
19. Vishnyakova P.A., Moiseev K.Y., Porseva V.V., et al. Somatostatinexpressing neurons in the tuberal region of rat hypothalamus during aging. J Evol Biochem Phys 2021; 57: 1480–1489. https://doi.org/10.1134/S0022093021060247
20. Lomax A.E., Sharkey K.A., Furness J.B. The participation of the sympathetic innervation of the gastrointestinal tract in disease states. Neurogastroenterol Motil. 2010; 22(1): 7–18. https://doi.org/10.1111/j.1365-2982.2009.01381.x
21. Palus K., Bulc M., Całka J. Changes in somatostatin-like immunoreactivity in the sympathetic neurons projecting to the prepyloric area of the porcine stomach induced by selected pathological conditions. Biomed Res Int. 2017: 9037476. https://doi.org/10.1155/2017/9037476
22. Van Op den Bosch J., Adriaensen D., Van Nassauw L., Timmermans J.P. The role(s) of somatostatin, structurally related peptides and somatostatin receptors in the gastrointestinal tract: a review. Regul Pept. 2009; 156(1–3): 1–8. https://doi.org/10.1016/j.regpep.2009.04.003
23. Gonkowski S., Rytel L. Somatostatin as an active substance in the mammalian enteric nervous system. Int J Mol Sci. 2019; 20(18): 4461. https://doi.org/10.3390/ijms20184461. PMID: 31510021
24. Ameri P., Ferone D. Diffuse endocrine system, neuroendocrine tumors and immunity: what’s new? Neuroendocrinology. 2012; 95(4): 267–276. https://doi.org/10.1159/000334612
Емануйлов Андрей Игоревич - канд. мед. наук, доцент кафедры анатомии человека
ул. Революционная, д. 5, г. Ярославль, 150000
Порсева Валентина Вячеславовна - д-р мед. наук, доцент кафедры патофизиологии
ул. Революционная, д. 5, г. Ярославль, 150000
Павлов Алексей Владимирович - д-р мед. наук, профессор, заведующий кафедрой гистологии
ул. Революционная, д. 5, г. Ярославль, 150000
Маслюков Петр Михайлович - д-р мед. наук, профессор, заведующий кафедрой нормальной физиологии
ул. Революционная, д. 5, г. Ярославль, 150000
Тел.: +7 (910) 972-88-61

|
1. Чек-лист ARRIVE 2.0 | |
| Тема | ||
| Тип | Прочее | |
Скачать
(115KB)
|
Метаданные ▾ | |
119048, г. Москва, ул. Трубецкая, д. 8, стр. 2
E-mail: sechenovmedj@staff.sechenov.ru

































